首页 > 心得体会 > 学习材料 / 正文
耐辐射奇球菌全局调控蛋白IrrE作为抗逆元件的机制及应用
2023-02-05 14:40:10 ℃黎 杰,陶惟一,李 霜
(1.南京工业大学 生物与制药工程学院 江苏 南京 211800;2.南京工业大学 食品与轻工学院 江苏 南京 211800)
微生物细胞工厂在生产过程中存在着一些不利环境因素,如,原料预处理带来的有毒抑制物、高盐、高温、高浓度的底物或产物带来的高渗透压以及溶剂等,从而影响细胞工厂的生产效能[1]。近年来,利用微生物抗逆元件构建高效细胞工厂成为研究热点。微生物抗逆元件主要包括调控细胞壁和细胞膜、DNA修复、氧化应激、相容性溶质、能量产生和信号转导的相关基因以及外排泵、热激蛋白和全局转录因子等[2-3]。其中全局转录调控工程(global transcription machinery engineering,gTME)成为改变复杂细胞表型的一种重要策略[4],该策略通过全局调控因子及其突变体调节宿主菌中数千个基因的表达,使得宿主菌激发出耐溶剂、耐酸、耐盐、耐热等优势表型[5-7]。这种针对多基因表达调控的代谢改造策略在一定程度上解决了载体构建、转化效率和筛选能力等方面的限制,成为提升细胞抗逆性能的重要策略之一。
IrrE也被称为PprI,是广泛存在于耐辐射极端微生物Deinococcus菌属中的特异性全局调控蛋白,通过参与调控DNA修复蛋白基因recA的表达,提升细胞耐受高剂量UV辐射、电离辐射及干旱等不良环境[8-9]。鉴于RecA蛋白及类RecA蛋白广泛存在于包括古细菌、真细菌、酵母、植物及动物等全部物种中,异源表达irrE是否能提升宿主对辐射的耐受性成为率先研究的对象。2003年,Gao等[10]在大肠杆菌中首次表达了来自耐辐射球菌的pprI基因(irrE),结果表明,组成型表达irrE基因使得宿主对高剂量γ线的耐受性提高了1.6倍,其作用机制为IrrE蛋白显著提高了宿主细胞中用于DNA修复的RecA蛋白表达量,并诱导提升过氧化氢酶(KatG)酶活,具有更好的清除胞内游离基的能力。2009年,Pan等[11]将irrE基因分别在大肠杆菌和油菜中进行了表达,发现IrrE蛋白可以赋予宿主对抗高盐、氧化压力、高渗透压以及高温等不良环境的能力。
此后,随着合成生物学技术的发展,有关IrrE蛋白对不同种属微生物的抗逆性能及作用机制的研究开始大量出现,IrrE开始成为合成生物学工具箱中一种重要的抗逆元件。笔者针对Deinococcus菌属的全局调控蛋白IrrE在异源宿主细胞的全局调控作用机制进行总结,并通过其在微生物细胞工厂中的应用案例展示IrrE作为抗逆元件的最新应用进展。
1956年,耐辐射极端微生物首次在高剂量γ线辐射后的肉罐头中被发现[12],并命名为耐辐射微球菌Micrococcusradiodurans,该菌株为革兰氏阳性球菌,菌落及液体培养物均为红色[13]。1981年,该菌株经系统鉴定后,被更名为耐辐射奇球菌Deinococcusradiodurans[14]。1999年,D.radioduransR1菌株的基因组完成了测序[15]。为更清楚地认识D.radioduransR1的电离辐射抗性,2002—2003年,先后有2个研究团队利用发生突变的电离辐射敏感菌株进行抗性基因解析,其结果均指向DR0167编码序列,随后命名为irrE基因或pprI基因,证明该基因编码蛋白可调控DNA修复功能蛋白基因recA的表达,对提高耐辐射抗性具有重要作用[16-17]。李新娜等[18]和张陈等[19]先后对耐辐射球菌DNA损伤修复重要功能蛋白进行了总结,陈震等[13]总结了耐辐射球菌全局调控蛋白IrrE的研究进展,对于国内同行开展相关研究提供了较为详尽的基础知识。
随着越来越多的该属其他菌株被分离鉴定和基因组测序,研究者发现,irrE基因广泛存在于该属菌株中,IrrE成为Deinococcus菌属的种属特异性调控蛋白。在已完成测序的Deinococcus菌株中共发现了7种不同来源的IrrE蛋白,这些IrrE蛋白的氨基酸序列同源性较高且具有相同的保守结构域,预示其功能上具有相似性。目前,已报道用于研究的3种IrrE蛋白分别来源于D.radioduransR1、D.deserti和D.gobiensisI-0这3个菌株;
采用序列比对软件分析表明,IrrE-D.radioduransR1与IrrE-D.gobiensisI-0、IrrE-D.deserti的同源性分别为65.22%和65.07%,IrrE-D.gobiensisI-0与IrrE-D.deserti同源性为73.02%(图1)。

图1 不同来源IrrE蛋白的序列同源性分析Fig.1 Sequence homology analysis of IrrE protein from different sources

图2 D. deserti全局调控蛋白IrrE的二级结构 (源于PDB数据库)Fig.2 Structure of global regulator IrrE from D. deserti(Originated from PDB database)
关于IrrE蛋白的功能结构域及作用机制的研究当前尚未有明确结论。针对D.deserti来源的IrrE蛋白结构解析发现N端(锌肽酶结构域)和C端(GAF结构域)是关键功能结构域[20]。最近对于IrrE蛋白N端功能的研究取得了一定的进展。Blanchard等[26]解析了Deinococcus菌属中IrrE/DdrO控制的耐辐射性状的保守性和多样性,同时还表明,DdrO蛋白具有多个DNA特异性结合位点。DNA损伤修复可以通过IrrE和DdrO共同调节[22,27-28]。在辐射应激作用下,胞内游离锌含量增加,刺激IrrE作为金属蛋白酶对阻遏蛋白DdrO的切割[29],解除对DNA损伤修复相关功能基因的抑制作用,导致与DNA和RNA代谢相关的多个基因(ddrC、ddrD、pprA、recA、ssb、ssb、uvrD、ddrF等)转录表达[30]。IrrE对DdrO特定结构域位点的切割,使得DdrO结构发生了动态平衡转换,脱离了DNA结合位点,解除相关基因的阻遏作用[31]。而针对D.radiodurans来源的IrrE研究表明,THT结构域的突变也能导致对辐射等不良环境敏感,推测THT结构域对DNA修复也有重要影响。2015年,Zhang等[32]将2个不同种(D.radiodurans和D.gobiensis)来源的irrE进行了突变和回补实验,有助于明确THT结构域的关键功能位点。D.gobiensisⅠ-0菌株比D.radiodurans菌株具有更好的耐辐射性能,有趣的是,当针对D.radiodurans的ΔirrE菌株进行回补实验时,来源于D.gobiensis的IrrE相较于出发菌株显示了更高的耐辐射活性,表明不同来源的IrrE具有活性上的差异。将来源于D.radiodurans的IrrE的THT结构域进行单点突变A184S,突变体表现出活性增加,而针对D.gobiensis的IrrE同位点的突变(S131A)却导致了活性的降低,推测THT结构域的丝氨酸残基可能是重要作用位点。
因此,目前对于IrrE结构的研究相对较为详细,但对于作用机制的研究还有所欠缺,主要的原因可能是IrrE结构的特殊性赋予了其许多复杂的调控功能,导致研究受到阻力。而当前研究部分表明,IrrE的3个结构域都参与了调控作用,有基因与基因间的调控,基因与蛋白间的调控或蛋白与蛋白间的调控。而最近研究较多的是IrrE和DdrO的调节,可以影响众多基因的转录表达,提高菌体耐辐射性能。但许多IrrE作用机制还需要进一步研究,需要最大程度地挖掘IrrE的潜在价值。
作为Deinococcus菌属的种属特异性全局调控蛋白IrrE,是否能在其他非亲缘种属菌株中展现全局调控能力并提升细胞对环境压力的耐受性,是科研工作者最为关注的内容。来源于D.radioduransR1的IrrE是目前最为广泛使用的转录调控蛋白,而大肠杆菌作为模式菌株,在研究全局转录调控蛋白IrrE的作用机制中最为详尽,多篇研究论文都证实了IrrE蛋白在大肠杆菌中具有全局调控能力,如图3所示。

GadE—可结合HTH结构域的调控因子;
GadX—参与DNA转录调节因子;
PurR—嘌呤核苷酸合成阻遏物;
AppY—含有ARAC型DNA结合转录激活因子;
GadW—ARAC型调节因子;
YhiF—可结合HTH结构域转录因子;
RpoS—σs;
BetI、CynR、MhpR、KdgR—转录调节因子;
PrpR—含有AAA型ATP酶结构域和DNA结合结构域的转录调节因子;
TdcA—tdc操纵子的转录激活因子,TdcR—DNA结合转录激活因子;
appBC—细胞色素bd型喹啉氧化酶基因;
hyaABCDEF—氢化酶基因簇;
有机溶剂耐受性相关基因(iraD、iraM)—RpoS蛋白水解抑制基因;
otsAB—海藻糖合成基因;
treBC—海藻糖转运及降解基因;
gldA、glpA、glpK—甘油降解基因;
氮代谢及转运相关基因narK—亚硝酸盐外排蛋白基因;
narGHIJ—硝酸还原酶有关基因;
nirBD—亚硝酸还原酶 (NADH)基因;
nirC—亚硝酸转运蛋白基因;
tdcB—苏氨酸脱水酶基因;
tdcC—氨基酸渗透基因;
tdcD—乙酸激酶基因;
tdcE—丙酮酸-甲酸裂解酶基因;
tdcF—翻译起始抑制基因;
tdcG—L-丝氨酸脱氨酶基因;
prpB—PEP磷酸变位酶基因;
prpC—柠檬酸合酶基因;
prpD—参与丙酸分解代谢的未表征蛋白质基因;
prpE—酰基辅酶A合成酶/AMP-(脂肪)酸连接酶基因;
betA—胆碱脱氢酶和相关黄素蛋白基因;
betB—NAD依赖性醛脱氢酶基因;
betT—胆碱-甘氨酸甜菜碱转运蛋白基因;
dctA—C4-二羧酸、乳清酸和柠檬酸转运蛋白基因;bfr—细菌铁蛋白(细胞色素b1)基因;
feoAB—亚铁转运蛋白基因;
fiu—儿茶酸铁载体受体/预测的铁外膜转运蛋白基因;
cirA—儿茶酸铁外膜转运蛋白基因;
fhuE—铁红酵母酸外膜转运蛋白基因;
fecABER—柠檬酸铁转运相关蛋白基因;
fecl—RNA聚合酶σ19因子基因;
fes—肠杆菌素/铁肠杆菌素酯酶基因;
fhuF—参与异羟肟酸铁转运的三价铁还原酶基因;
fepBC—铁肠杆菌素转运相关基因;
fhuA—铬铁外膜转运蛋白基因;kdgK—糖激酶基因;
kdgT—2-酮-3-脱氧葡萄糖酸渗透基因;
kdulD—葡萄糖代谢相关基因;
yjgK—β-半乳糖苷酶β亚基基因;
mhpA—3-羟苯基丙酸羟化酶基因;
mhpD—2-酮-4-戊烯酸水合酶基因;
mhpE—4-羟基-2-氧代戊酸醛缩酶基因;
mhpT—3-羟苯基丙酸转运蛋白基因;
cynS—氰酸裂解酶基因;
cynT—碳酸酐酶基因;
purB、purC、purEK、purF、purHD、purL、purMN—核苷酸合成过程有关基因;
pyrD—二氢乳清酸脱氢酶基因;
glyA—丝氨酸羟甲基转移酶基因;
gadYXW—酸适应性调控基因;
hdeAB—周质酸应激伴侣蛋白基因;
hdeD—抗酸膜蛋白基因;
yhiD—假定的参与耐酸的一种膜蛋白基因;
slp—一种外膜脂蛋白基因图3 IrrE对大肠杆菌的全局调控作用机制[35-36,38]Fig.3 The global regulatory mechanism of IrrE on Escherichia coli[35-36,38]
潘婕[33]和周正富[34]率先开展了基于转录组和蛋白组的分析,研究了IrrE对大肠杆菌的影响。Zhou等[35]研究了IrrE蛋白对E.coliJM109在非压力环境下的转录组和蛋白组影响;
结果表明,IrrE蛋白对大肠杆菌的1个EvgSA(具有传感和调节作用的两种蛋白)双组分系统、3个主调控因子GadE、GadX和PurR以及10个转录因子(AppY、GadW、YhiF、AsnC、BetI、CynR、MhpR、PrpR、TdcA和KdgR)起到了调控作用,可以导致数百个基因表达差异,包括海藻糖合成、核苷酸合成、碳源代谢、氨基酸代谢等代谢途径以及细胞运输系统。为了进一步探索IrrE对大肠杆菌耐盐胁迫的全局调控作用机制,Zhao等[36]考察了IrrE蛋白对于宿主(E.coliJM109)在受到盐胁迫(1 mol/L NaCl)条件的基因表达差异,得到了相对直观和明确的结果。IrrE蛋白可以使宿主的100~300个左右的基因差异表达,影响宿主胞内海藻糖、甘油、核酸、碳源、氨基酸、抗酸性,以及一些依赖于转录因子RpoS(σS)的相关基因,如,海藻糖合成基因otsAB、抗酸基因gadABC和uspB、高渗透压和氧化响应基因katE和osmBC等,导致胞内相容性物质如海藻糖和甘油浓度的显著上升。而进一步分析表明,IrrE并没有影响rpoS的转录,只是过表达一些基因阻止了RpoS蛋白的降解,使得转录因子RpoS蛋白得到了积累,从而影响了众多基因的表达。这些研究结果表明,IrrE蛋白可以在大肠杆菌中发挥全局调控功能。
清华大学林章凛团队针对IrrE蛋白开展了突变和定向进化研究,进一步拓展了IrrE的适用性。Chen等[37]首次构建了IrrE的突变体文库,以E.coliDH5α为宿主,筛选突变体对乙醇、丁醇和乙酸的选择耐受性,发现部分IrrE突变体可以让宿主的生物量增加2~50倍,同时大幅降低宿主的胞内活性氧(ROS)。针对突变位点的分析表明,这些突变位点分布于IrrE的所有结构域,并没有显著的结构特征。此后,Chen等[38]进一步分析了野生型IrrE和突变体对宿主的转录组和蛋白组的影响;
结果表明IrrE突变体对宿主的转录组和蛋白组的影响更显著,总计有1 196个基因在转录水平上有显著改变,上调和下调的蛋白斑点数均达到100多个,特别是硝酸盐、色氨酸、Fe离子和甘油以及与氧化磷酸化和非编码RNA相关的基因在转录水平上显著上调或下调。Wang等[39]将IrrE突变体用于提升大肠杆菌对木质纤维素水解液抑制物的耐受性,在体积分数为0.2%糠醛的条件下,IrrE突变体的重组菌生物量是其对照组的4~16倍,且对5-羟甲基糠醛、香兰醛以及纤维素水解液均具有显著的耐受性。此后,骆健美等[40]将irrE基因及其突变体在E.coliBL21(DE3)中进行异源表达,也获得类似的结果,IrrE及其突变体能够不同程度提高宿主菌在各种胁迫条件下(3 mol/L山梨醇、pH 2和pH 12、体积分数为10%甲醇、体积分数为12%乙醇和体积分数为1% H2O2)的存活能力。
这些研究结果均表明,IrrE在大肠杆菌中具有很好的全局调控能力,引起众多基因的差异表达,激发宿主耐受各种极端环境。这些引起宿主的差异变化主要表现在抗氧化还原,提高有机溶剂耐受性、抗渗透压、耐酸、氮代谢及转运、碳代谢、氨基酸代谢、Fe离子转运、核苷酸合成等。而且,通过突变或定向进化筛选,可以获得应对不同极端环境的IrrE。因此,IrrE比其他全局转录调控因子具有更广泛的应用价值,可应用于耐受多种极端环境。
在工业微生物的发酵体系以及发酵进程中,高浓度底物/产物导致的高渗透压、代谢产物或副产物积累导致的酸、碱、溶剂等容易导致微生物处于不利于生长或代谢产物积累的环境,进而限制了产物浓度、转化率及生产速率等指标的提升。目前,广泛地将耐辐射奇球菌的IrrE蛋白作为抗逆元件应用到含有醇、酸、渗透压、电场等环境,提高宿主菌在各种环境中的耐受性,其中涉及应用的宿主菌有革兰氏阳性菌(乳酸菌、节杆菌)、革兰氏阴性菌(大肠杆菌、运动单胞菌、鞘氨醇单胞菌、假单胞菌等)以及酿酒酵母等,这是近年来将IrrE蛋白从原始菌株到各种宿主菌作为抗逆元件应用的热门研究方向。
3.1 irrE在燃料乙醇生产菌株中的应用
利用木质纤维素原料生产燃料乙醇是生物能源的重要研究课题。木质纤维素原料预处理产生的糠醛类抑制物和高浓度的乙醇及副产物等会对生产菌产生损伤,从而影响乙醇的生产速率和转化率[41]。全局调控因子IrrE及其突变体在各种乙醇生产菌株(大肠杆菌、运动单胞菌、酿酒酵母)中均可激发宿主菌获得耐受胁迫环境的新表型,如图4所示。
E.coliMG1655是研究有机酸和乙醇代谢的模式菌株。Ma等[42]发现在MG1655中表达irrE基因后,宿主菌对高渗透压(250 g/L的葡萄糖/木糖)和酸胁迫(pH 2)的耐受性显著增强,且宿主菌的乙醇积累能力也显著提升;
以100 g/L葡萄糖或木糖为碳源时,宿主菌的乙醇积累量分别为39.63和34.53 g/L,相比对照菌株产量分别提高了14.7%和26.3%;
在发酵过程中,宿主菌中与乙醇合成途径相关的关键酶PDC(丙酮酸脱羧酶)和ADHB(乙醇脱氢酶)的活性均持续性显著高于对照组的活性。
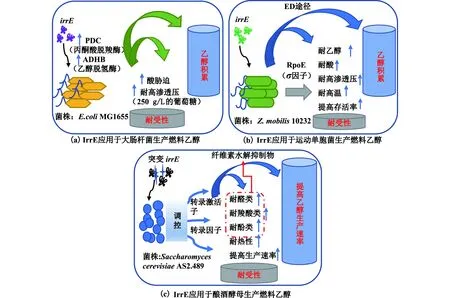
图4 IrrE作为抗逆元件在乙醇生产菌株中的应用Fig.4 Application of IrrE as tolerance element in ethanol production strains
运动单胞菌Zymomonasmobilis可通过其特有的ED(2-酮-3-脱氧-6-磷酸葡糖酸裂解途径)途径高效积累乙醇,是具有工业应用潜力的乙醇生产菌株。Zhang等[43]在Z.mobilis10232中表达了irrE基因,发现重组菌提高了对乙醇、酸、高渗透压及高温等不良环境的耐受性和存活率,尤其值得关注的是,在初始pH 3.8和体积分数为12%的乙醇环境下,重组菌乙醇积累量分别为18.8和14.59 g/L,而对照组的乙醇积累量分别为14.2和8.75 g/L,且重组菌中转录因子RpoE(一种σ因子)的转录水平分别较对照组提升了8.2倍和6倍,提示该菌株中与耐受性相关基因可能受到转录因子RpoE的调控。
酿酒酵母是当前燃料乙醇产业的主要生产菌株。2018年,Luo等[44]在SaccharomycescerevisiaeAS2.489中表达irrE突变体获得了对糠醛的耐受性,同时对5-羟甲基糠醛、甲酸、乙酸等木质纤维素预处理体系抑制物都具有抗性的重组菌;
在含2 g/L糠醛的合成培养基(200 g/L葡萄糖)中,重组菌的乙醇产量(88 g/L)没有显著差异,但生产速率提升了37%。2020年,Wang等[45]发现IrrE突变体可以将S.cerevisiaeBY4742在木质纤维素水解液抑制物及高温(42 ℃)等不良环境下的乙醇发酵速率提升3倍以上;
并进行转录组分析发现,在酿酒酵母胞内IrrE作为全局调控因子,可以调控转录激活子及转录因子的转录水平,从而实现在基因组尺度上对转录进行扰动。Helalat等[46]在野生型酵母S.cerevisiaeINS中异源表达pprI(irrE)获得的重组菌也可以提高细胞对盐(70 g/L)、乙醇(体积分数为11%)、丁醇(体积分数为1%)的耐受性。
这些研究结果表明,全局调控因子IrrE可以激发各种燃料乙醇生产菌株的抗逆能力,提高菌株的生产性能。目前做过相关尝试的菌株主要有大肠杆菌、运动单胞菌、酿酒酵母。IrrE在这些菌株中都表现出了较好的全局调控能力,激发了菌株耐受渗透压、乙醇、高温和纤维素水解抑制物等不利环境因素的能力,提高了菌体的存活率和乙醇的生产性能。因此,IrrE调控蛋白作为抗逆元件,可以有效地突破菌体自身极限,提高菌体生产性能。
3.2 irrE在其他微生物细胞工厂中的应用
近年来,来源于D.radiodurans的全局调控蛋白IrrE在利用微生物发酵产有机酸、生物聚合物、甾体转化以及燃料电池、生物修复等领域进行了应用尝试[47,48-54],如表1所示。从应用中发现,野生型IrrE在革兰氏阳性菌(乳球菌、节杆菌)和革兰氏阴性菌(鞘氨醇单胞菌、假单胞菌、大肠杆菌)中都能提高宿主细胞对特定环境的耐受性,大幅提升生产效率和产量;
且相应的研究表明IrrE可扰动宿主中多基因表达差异,从而影响宿主表型,赋予宿主更加优异的生产性能。
在有机酸发酵中,酸、渗透压、盐以及氧化压力等对生产菌株的胁迫是一个比较常见的问题。Dong等[47]在乳酸菌中异源表达irrE,发现可以提高菌株耐渗透压和抗氧化压力,提高高盐胁迫下的乳酸产量。朱兴贵等[49]、Zhang等[50]发现IrrE可提升大肠杆菌耐盐性能和发酵产琥珀酸(提升1.24倍)的性能,使得利用海水代替淡水发酵产琥珀酸成为可能。

表1 IrrE在微生物工厂中的应用Table 1 Application of IrrE in microbial cell factoryies
在甾体生物转化领域,添加溶剂(乙醇)可以提高底物溶解度,有助于提升转化效率。Song等[51]、Luo等[52]以简单节杆菌A.simplexCPCC 140451转化醋酸可的松生成醋酸泼尼松为模式体系,发现表达irrE后的重组菌可提升对乙醇的耐受性,在高浓度乙醇作为助溶剂时,其产物生成速率和最终产量都有显著提高,通过对irrE表达水平的优化,可以获得耐胁迫能力最佳菌株,类固醇产量提高了261.8%。最为有趣的是,irrE基因也在A.globiformisATCC 8010和E.coli中表达,结果发现这3个重组菌对有机溶剂和氧化压力都提升了耐受性;
但是,只在E.coli中表现出提升了对酸/碱休克的耐受性,只在A.simplex中表现出对热休克更好的耐受性。这些研究表明IrrE作为抗逆元件,其表达量会影响宿主的抗逆性能及生产性能,在不同宿主或应对不同压力环境时也会有差异性。
鞘氨醇单胞菌Sphingomonas是威兰胶、结冷胶、定优胶等生物聚合物的主要生产菌株。最近,Liu等[48]在威兰胶生产菌株中表达了irrE,转录组分析发现扰动了1 000多个基因(双组分系统、细菌趋化性、鞭毛组装、生物膜形成和细胞周期)表达,使得重组菌可以耐热、耐pH等胁迫压力,从而降低生产能耗和成本。
假单胞菌属微生物代谢能力强,在降解有毒化合物方面具有广泛应用。Zhou等[54]发现表达irrE可提升恶臭假单胞菌S16的耐酸性能,使得其可以在低pH的条件下降解污染物(苯甲酸或尼古丁)。Luo等[53]将irrE引入铜绿假单胞菌PAO1菌株,发现IrrE对宿主的转录组可产生扰动,菌株的产电性能提高71%。
上述研究表明,全局调控蛋白IrrE及其突变体已经应用到了各领域中,可以普遍地和多种微生物中的基因建立联系,引起转录扰动,激发宿主菌耐受(渗透压、盐、酸、热、氧化压力、有机溶剂等)不利环境因素的能力。其中,IrrE调控蛋白对宿主的碳氮源代谢、能量代谢、细胞膜形成、甘油、海藻糖等相关途径的基因扰动最受关注,但也将视野拓展到了次级代谢途径及QS调控因子,例如,吩嗪生物合成、细菌趋化性、鞭毛组装等基因[47-48,53]。未来,随着IrrE应用领域的扩宽,新的应用宿主和表型会更加丰富。
由于极端微生物在长期的自然胁迫条件下进化出了一套特殊、高效的胁迫响应机制,因此其往往成为挖掘抗逆元件的资源库。来源于耐辐射极端微生物Deinococcus菌属的全局转录调控蛋白IrrE大约已有20多年的研究历程,其应用性范围不再局限于提高菌体的耐辐射能力。从2009年发现IrrE蛋白可以赋予宿主抗逆表型(抗高盐、抗氧化压力、抗高渗透压以及高温)开始,IrrE蛋白成为一种可跨越种属的限制与广泛宿主建立起全基因组尺度的全局转录调控因子,通过复杂的转录组水平上的调控差异,激发出新的表型。例如,IrrE蛋白可以激发各类(乳酸菌、节杆菌、大肠杆菌、运动单胞菌、鞘氨醇单胞菌、假单胞菌和酿酒酵母等)菌体对极端环境的耐受能力,提高细胞自身的生产性能。因此,IrrE蛋白用于提高菌体耐辐射能力的同时,也可以打破物种的限制,用于提高各类菌体耐极端环境(抗高盐、抗氧化压力、抗高渗透压和抗高温等)的能力,提高菌体的生产性能。而且,由于IrrE蛋白比其他全局转录调控因子具有更广泛的应用范围,因此,IrrE蛋白作为一种抗逆元件是近几年的研究热门。
耐辐射极端微生物Deinococcus菌属的菌株在分类学上已达20余种,作为该属菌株中普遍存在的irrE基因,这些不同来源的野生型irrE基因是极其丰富的资源。在针对特定宿主及抗逆表型的应用研究中发现,不同来源irrE基因对宿主的作用效果存在差异,而且IrrE蛋白的表达量也对宿主表型产生影响。因此,可以通过筛选IrrE蛋白突变体赋予宿主细胞对抗特定环境的能力。同时,如果野生型IrrE蛋白不能满足宿主的抗逆性能需求,通过定向进化方式获得“适配”的突变体可以进一步丰富抗逆元件“工具箱”。
因此,总结前期的一些研究结果可以发现,尽管来源于耐辐射极端微生物的抗逆元件IrrE蛋白在不同物种中的作用机制还不够清晰,但却不影响人们对它的应用和尝试。而且,随着IrrE蛋白作为抗逆元件在各类细胞中的广泛应用尝试,未来可能会出现具有针对性的或多样化的IrrE抗逆元件“工具箱”。IrrE蛋白的广泛适用范围和可以激发宿主许多优良的抗逆表型使其在合成生物学领域具有丰富的应用前景。
猜你喜欢 耐受性宿主结构域 细菌四类胞外感觉结构域的概述湖北农业科学(2022年11期)2022-07-18UBR5突变与淋巴瘤B细胞成熟实用肿瘤学杂志(2020年4期)2020-12-08龟鳖类不可能是新冠病毒的中间宿主当代水产(2020年3期)2020-06-15黄鳝对常用消毒药物的耐受性反应研究现代农业科技(2016年20期)2016-12-20抓住自然宿主小星星·阅读100分(高年级)(2015年11期)2015-11-28绦虫大战,争夺宿主控制权大自然探索(2015年11期)2015-09-10护理干预对ICU患者肠内营养耐受性影响的探讨现代养生·下半月(2015年6期)2015-09-07小儿胃镜检查时耐受性与安全性的临床研究现代养生·下半月(2015年2期)2015-06-15人乳头瘤病毒感染与宿主免疫机制医学研究杂志(2015年12期)2015-06-10DEP结构域的功能研究进展医学综述(2011年12期)2011-12-09猜你喜欢
- 2024-01-20 有关于第五次全国经济普查统计重点业务综合培训大会上讲话(完整文档)
- 2024-01-20 “严纪律、转作风、保安全、树形象”专题学习教育活动通知(完整文档)
- 2024-01-20 2024XX区住房城乡建设工作情况汇报
- 2024-01-20 2024高校思政教育交流材料:善用反腐败斗争这堂“大思政课”(精选文档)
- 2024-01-20 2024年主题教育专题党课辅导报告,(4)
- 2024-01-20 关于赴某地学习考察地方立法工作情况报告(范文推荐)
- 2024-01-20 2024年度关于增强党建带团建工作实效对策与建议(精选文档)
- 2024-01-20 教师演讲稿:春风化雨育桃李,,潜心耕耘满芬芳(全文)
- 2024-01-20 主题教育第二阶段来了
- 2024-01-20 2024年度关于到信访局实践锻炼个人总结【完整版】
- 搜索
-
- 打赌输了任人处理作文1000字7篇 05-12
- 当代大学生在全面建设社会主义现代化强 05-12
- 全面建成社会主义现代化强国的战略安排 03-10
- 个人廉洁自律方面存在的问题及整改措施 05-12
- 谈谈青年大学生在中国式现代化征程上的 05-12
- 2022年党支部第一议题会议记录(全文完 11-02
- 作为青年大学生如何肩负时代责任6篇 05-12
- 村党组织建设现状及工作亮点存在问题与 05-12
- 全面从严治党,自我革命重要论述研讨会 05-12
- 产业工人队伍建设改革(全文完整) 10-31
- 11-25国庆70周年庆典晚会 庆典晚会串词
- 11-25办公室礼仪的十大原则 浅谈办公室的电话礼仪
- 01-17用心灵轻轻地歌唱_心灵的歌唱
- 01-17也许你不是我一生的唯一|也许不是我
- 01-17爱了,请珍惜;不爱,趁早放手|爱就珍惜不爱就放手
- 01-17岁月带走的是记忆,但回忆会越来越清晰|有趣又有深意的句子
- 01-17曾经的美好只是曾经,我只想珍惜身边的人|我只想珍惜你
- 01-18从容不惊 [学会笑眼去看世界,不惊不乍,淡定从容]
- 02-03当代大学生学习态度调查报告
- 02-03常用护患英语会话
- 标签列表
