首页 > 心得体会 > 学习材料 / 正文
姬松茸菌褶、担孢子及菌丝体微观形态观察
2023-03-04 16:15:18 ℃周 宇,黄在兴,鲍兴禄,李 晶,刘朋虎*
姬松茸菌褶、担孢子及菌丝体微观形态观察
周 宇1,2,黄在兴3,鲍兴禄4,李 晶2,刘朋虎2*
1. 福建农林大学园艺学院,福建福州 350002;
2. 福建农林大学国家菌草工程技术研究中心,福建福州 350002;
3. 广西大学农学院应用微生物研究所/广西大学食用菌研究所,广西南宁 530005;
4. 福建省武夷山市吴屯乡乡村振兴发展中心,福建武夷山 354309
姬松茸()不仅营养丰富还具有很高的药用价值,是一种名贵的食药用菌,其产业发展前景广阔。但由于姬松茸种质资源匮乏,对姬松茸的遗传背景认识不足,阻碍了其杂交育种技术的发展,现有的种质资源难以满足实际生产需要。本研究以姬松茸J1菌株为实验材料,通过对姬松茸生殖相关结构进行微观形态观察,探究姬松茸相关的生物学特性。利用扫描电子显微镜和荧光显微镜观察姬松茸不同生长发育时期菌褶、担孢子和菌丝体微观形态特征。结果表明:桑葚期菌褶和菌髓处于发育阶段,菌褶片状物不明显;
珊瑚期后能明显观察到菌褶片状物,相互平行排列,由菌髓和其两侧的子实层组成,排列紧密,厚度均匀,且着生少量担子,但菌髓呈不规则排列;
成形期菌褶更为清晰、排列规整,内菌幕未破裂,菌褶间隙包含表面凸起排列的担孢子和杂乱的导管状细胞,导管状细胞紧密关联;
成形期菌褶担子较为饱满,有褶皱,担子上着生有3个孢子梗或4个孢子梗,各担孢子之间有细小的连接丝。通过DAPI对担孢子核数进行染色观察,发现姬松茸大多数担孢子属于双核孢子,少量属于单核和无核孢子;
通过DAPI对菌丝体细胞核进行染色观察,发现姬松茸菌丝体属于多核,有隔膜,但未见锁状联合。本研究明确了姬松茸不同生长发育时期的菌褶、担子、担孢子以及菌丝体的微观形态特征,丰富了姬松茸的生物学研究和生活史过程认识,为姬松茸杂交育种、担孢子交配型、遗传转化体系等研究奠定了基础。
姬松茸;
菌褶;
担孢子;
细胞核
姬松茸()又名巴西蘑菇、阳光蘑菇、小松菇、巴氏蘑菇等,不仅味道鲜美,营养丰富,还具有多种药用价值,在降血脂、抗糖尿病、抗炎、免疫调节以及抗癌等方面表现出较好的抑制或治疗作用[1-5]。姬松茸在美食、保健、医学和药学界受到广泛关注。近年来,经过引种栽培,姬松茸已在中国、日本、美国、巴西等国家大面积推广栽培,其产业发展总体保持较好势头,但同时也面临产量不稳定、重金属富集能力强、种质资源匮乏等问题,阻碍了该产业的可持续发展[6]。
优良菌种的选育是解决上述问题的有效手段[6-7]。然而,目前对姬松茸交配型、生活史过程相关场所的显微变化等育种相关的基本生物学信息的了解十分有限,限制了杂交育种技术在姬松茸优良菌种选育上的应用。菌褶作为食用菌担孢子的发育场所,功能性孢子的形成对于食用菌生殖和育种至关重要;
其次,菌褶在蘑菇分类与生活史研究中也具有重要作用。目前,已有众多学者关于食用菌菌丝体、担孢子扫描电镜样品制备方法的探索,对于不同的食用菌部位和不同的观察要求,所参考的方法不尽相同[8-10]。但目前尚无姬松茸不同生长发育阶段菌褶、担子、担孢子以及菌丝体等的显微结构变化的相关报道,本研究运用扫描电子显微和荧光显微观察技术验证并完善姬松茸的不同生长发育阶段的形态特征,旨在丰富食用菌育种相关的基础生物学研究,以期为遗传育种、担孢子交配型、遗传转化体系等研究提供理论依据。
1.1 材料
1.1.1 供试菌株 供试菌株为姬松茸J1,菌株保藏于国家菌草工程技术研究中心菌种保种室。
1.1.2 培养基 复壮培养基:马铃薯230 g,蔗糖20 g,磷酸二氢钾2 g,硫酸镁0.5 g,维生素B1 10 mg,琼脂20 g/L,加蒸馏水定容至1 L,pH为7。
母种培养基:麦粒100%。
发酵料配方:稻草35.70%,牛粪14.29%,麸皮13.29%,谷壳35.70%,KH2PO40.02%,CaCO31%。
1.1.3 试剂与仪器 5%戊二醛,0.2 mol/L磷酸缓冲溶液(pH 7.4),1%锇酸,10 μg/mL DAPI染色液均购于北京兰杰柯科技有限公司;
日立(HITACHI)HCP-2型临界点干燥仪,日本EIKO IB-5型离子镀膜仪,日本JEOL JSM-6380LV扫描电子显微镜和日本Nikon NI-U荧光显微镜。
1.2 方法
1.2.1 子实体形态观察 姬松茸是一类具有菌褶的伞状真菌。为了下一步的菌褶扫描电子显微镜观察实验,本研究选取桑葚期、珊瑚期、成形期和成熟期4个不同发育时期为观察对象[11]。不同时期子实体采集标准见图1。桑葚期能分辨出为伞形,子实体棕褐色;
珊瑚期菌盖开始撑开,能看见内菌幕生长;
成形期内菌幕形成,菌柄伸长,色泽圆润洁白;
成熟期内菌幕逐渐消失。
1.2.2 菌褶扫描电子显微镜观察 将采集的新鲜子实体菌褶部位进行预处理,处理方法参照赵建霞等[12]的方法略有改动。取不同时期的菌褶放入离心管,用5%戍二醛固定样品4 h;
弃预固定液,使用0.2 mol/L磷酸缓冲液(pH 7.4)漂洗样品,清洗3次,每次10~15 min;
再使用1%锇酸固定4 h(通风橱内进行);
然后使用0.2 mol/L磷酸缓冲液(pH 7.4)再漂洗;
使用50%、70%、80%、90%、100%酒精逐级脱水,每遍间隔10~15 min,其中100%酒精置换3次;
使用环氧丙烷作为中间溶剂转换2次;
采用临界干燥仪干燥,使样品表面不受或少受表面张力影响的条件下除去乙醇;
处理好后用离子镀膜仪进行喷金。最后利用扫描电子显微镜观察。
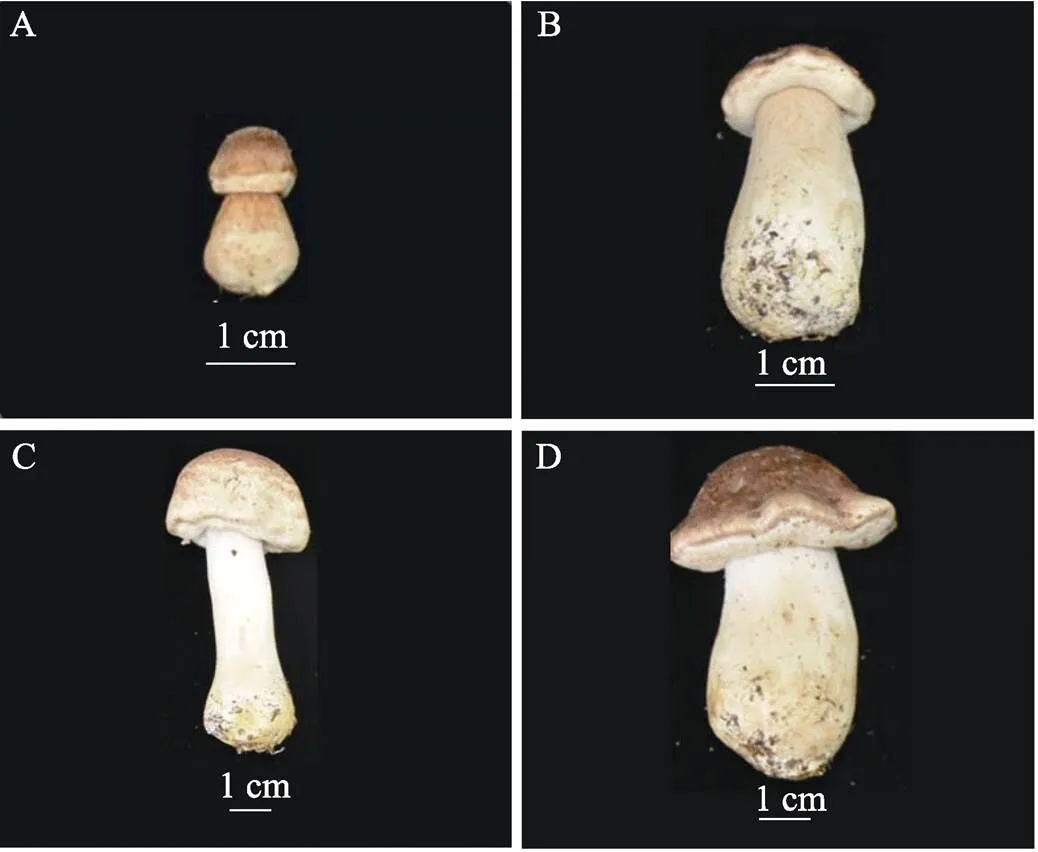
A:桑葚期;
B:珊瑚期;
C:成形期;
D:成熟期。
1.2.3 菌褶担孢子、担孢子和菌丝细胞核荧光显微镜观察 将刚采集回来的成形期子实体,用干净的镊子轻轻将菌褶分离,染色后于荧光显微镜下观察担孢子细胞核。担孢子细胞核的观察通过孢子印法收集孢子,对孢子进行稀释后染色观察。菌丝体细胞核的观察,利用复壮培养基培养菌丝,将配置好的培养基高温灭菌后,在超净工作台上把培养基倒入平板,等待至凝固,使用打孔器接种姬松茸菌丝到平板中央,并把盖玻片45°斜插入培养基上,待菌丝长至盖玻片2/3时,取出盖玻片进行染色观察。
荧光显微观察参照胡晓棣等[13]的染色方法略有改动,以上样品统一使用DAPI进行染色观察,将10 μg/mL DAPI利用PBS缓冲液(pH 6.8, 0.1 mol/L)进行稀释,DAPI最终染色浓度为5 μg/mL。将上述准备好的样品,利用DAPI(5 μg/mL)对样品进行避光染色5 min,最后置于荧光显微镜激发光波长340 nm与发射光波长488 nm下观察,激发出蓝色荧光。
2.1 姬松茸菌褶形态变化
对姬松茸桑葚期菌褶观察发现,子实层体呈弯曲状,菌褶处于发育阶段,观察到明显菌褶形态,菌髓还处在发育阶段(图2A)。进一步对图2A菌褶处放大观察,发现子实体基部细胞还处于生长发育阶段,菌褶上的担子细胞已有雏形初现,极少部分担子细胞末端开始膨大(图2B)。对姬松茸珊瑚期菌褶观察发现,菌褶排列紧密厚度均等,具有一定的规则性,且能明显看到片状物。图2C以一条150°斜线为界限,角度大于150°的部分就是菌褶(图2C-a),小于150°的部分为菌褶的基部(图2C-b),可以看到菌褶是由菌髓和其两侧的子实层组成,子实层上着生担子。相较于排列整齐的担子,菌髓呈不规则排列。对图2C菌褶部分依次进行放大观察(图2D~图2F),结果发现,菌褶中的基部细胞和担子细胞已有明显的分化趋势。

A、B:桑葚期菌褶;
C~F:珊瑚期菌褶, a为菌褶,b为菌褶基部。
2.2 姬松茸菌褶间隙导管状细胞
姬松茸菌褶是菌盖子实层体的皱褶部分,由菌褶原发育而成的片状结构。从横切面看,每个菌褶的两侧有子实层。姬松茸菌褶子实层的下层有一圈子实层基,即菌髓。担子着生于子实层,对姬松茸菌褶间隙依次放大观察发现,菌褶间隙包含表面凸起排列的担孢子和杂乱的导管状细胞(图3A,图3B)。此外,每片菌褶呈相互平行排布,且菌髓内细胞组织的排列与担子大致是垂直关系,这可能与固定担孢子形态和运输营养物质有关。桑葚期菌褶基部里的导管状细胞紧密关联,推测这种导管状细胞具有支撑和输送作用(图3C,图3D)。

A、B:菌褶与菌褶之间缝隙图;
C、D:菌褶内部导管状细胞。
2.3 姬松茸担子形态
姬松茸担子之间有细小的连接丝,担子较为饱满,未发现着生孢子梗(图4A~图4D);
通过进一步放大观察,可以观察到3个孢子梗或4个孢子梗,同时还观察到担子有褶皱(图4E,图4F)。从2个担子顶端的孢子梗可以看到,褶皱的的孢子梗开始膨胀,这可以说明姬松茸子实体发育后子实层裸露,其上的担子由饱满圆滑形状转变成褶皱蜂窝状,顶端的孢子梗膨胀有释放担孢子的趋势。
2.4 成形期菌褶担孢子荧光显微观察
姬松茸成形期菌褶的表面着生大量担孢子(图5),能明显观察到双核(图5A,图5B)。收集该时期弹射的孢子,对其进行荧光条件下显微观察(图5C,图5D),进一步确认担孢子大多属于双核担孢子(图5D),也有少量单核和无核(图5D),但担孢子的双核属于同核或者异核还需进一步研究确认。

A~D:成形期担子;
E~F:成形期担子着生3个或4个孢子,a和b为4个孢子梗,c为3个孢子梗。

A、B:菌褶碎片(小红圈代表双核孢子);
C、D:荧光观察图(红箭头指向双核孢子,白箭头指向单核孢子,黄箭头指向无核孢子)。Bars=100 mm。
2.5 菌丝细胞核观察
利用DAPI进行姬松茸菌丝染色观察,结果如图6所示,姬松茸细胞核颜色为蓝色,红色箭头所指的是菌丝细胞核,可以观察到姬松茸菌丝为多核,且有隔膜;
通过进一步观察,未发现锁状联合。

红色箭头指向细胞核;
蓝色箭头指向隔膜。Bars=100 mm。
本研究通过对姬松茸子实体不同发育时期的菌褶变化和担孢子发育过程进行显微观察,发现菌褶与担孢子存在发育不同步,菌褶为担孢子生长输送营养物质,并提供生长发育空间。子实体不同发育时期菌褶变化差异较大,桑葚期有菌褶雏形出现,该时期的菌褶还处于待发育状态,但能看见少量的担孢子,说明担孢子形成初期主要靠菌髓和子实层提供营养;
珊瑚期能明显看出菌褶片状物,此时的担孢子凸起并出现褶皱;
成形期担孢子上着生孢子梗,观察到担孢子上着生有3个或4个孢子梗,并观察到担孢子之间有丝状连接物,这与同属的双孢蘑菇观察到的结果较为一致[12]。而HERREIRA等[14]利用扫描电镜观察姬松茸担孢子,发现菌株CS2和CS7以2个孢子梗和3个孢子梗为主,菌株CS1、CS9和CS10以4个孢子梗为主,说明不同菌株间的孢子梗具有差异性。通过食用菌菌褶形态特征的扫描电镜观察,有助于开展食用菌孢子收集和孢子萌发的相关育种工作。许多研究者大都选择八成熟或成熟期的子实体进行单孢分离试验[15-17],而对于成形期的研究报道甚少。成熟期收集的孢子较易感染杂菌,选择成形期的孢子进行分离实验其活力是否可行,还需进一步实验验证。
解析食用菌不同发育时期菌褶的形态特征,对深入研究食用菌分子方面的机理具有一定参考价值。陈美元等[18]以双孢蘑菇菌褶为转化受体,运用农杆菌转化法将耐热相关基因028-1成功转入双孢蘑菇非耐热菌株8213。乔燕楠等[19]采用根癌农杆菌转化法,将双元载体pCghG转化双孢蘑菇不同组织,结果表明,菌褶部位转化率最高,达67.1%。喻义赣[20]利用香菇不同组织进行遗传转化试验,研究表明,幼嫩菌褶是农杆菌转化香菇的最适组织。食用菌菌褶部位被广泛运用于分子遗传转化,作为食用菌遗传转化体系的理想材料。目前,还未建立姬松茸稳定有效的遗传转化体系,姬松茸的相关分子研究受到一定制约,完善姬松茸的基本生物学研究是当下至关重要的一步。
此外,本研究还对姬松茸细胞核进行观察,结果表明姬松茸担孢子大多属于双核,菌丝体为多核,有隔膜,未见锁状联合。而杨利梅[21]对砖红线盖牛肝菌()进行显微观察发现,牛肝菌菌丝为双核,无锁状联合,这与本研究结果类似。而另有研究表明,刺芹侧耳()、凤尾菇()、灵芝()等食用菌均观察到细胞核和锁状联合[22-23]。这说明同一担子菌门不同属之间,菌丝体结构和细胞核数存在差异。本研究观察到担孢子双核和菌丝体多核对姬松茸的杂交育种工作提供理论基础。
(1)姬松茸子实体桑葚期菌褶还未发育健全,片状物不明显;
珊瑚期后能明显观察到菌褶片状物,能看见少部分担子;
成形期菌褶清晰排列规整,内菌幕未破裂。
(2)姬松茸菌褶间隙有大量导管状细胞,菌褶里面导管状细胞紧密关联,并起支撑和输送作用。
(3)姬松茸成形期菌褶着生担子,担子上着生有3个孢子梗或4个孢子梗;
姬松茸担孢子大多属于双核,菌丝体为多核,未见锁状联合。
[1] CAMPELO M D S, NETO J F C, LIMA A B N, NETO F C D C, NÁGILA M P S R. Polysaccharides and extracts fromMurill-a comprehensive review[J]. International Journal of Biological Macromolecules, 2021, 183: 1697-1714.
[2] KATSUYA T, HOICHI A, AKITOMO M, DAISUKE Y, KEN-ICHI I, YOSHIYUKI A, NAOHITO O. Outdoor-cultivated royal sun medicinal mushroomKA21 (Agaricomycetes) reduces anticancer medicine side effects[J]. International Journal of Medicinal Mushrooms, 2021, 22(1): 31-43.
[3] RUBEL R, SANTA H S D, SANTOS L, FERNANDES L, SOCCOL C. Immunomodulatory and antitumoral properties ofandmushrooms[J]. International Journal of Medicinal Mushrooms, 2018, 20(4): 393-403.
[4] 顾可飞, 邵 毅, 李晓贝, 范婷婷, 刘志敏, 黄柳娟, 王红梅, 周昌艳. 松茸与姬松茸营养成分差异性研究[J]. 农产品质量与安全. 2017(1): 73-78.
GU K F, SHAO Y, LI X B, FAN T T, LIU Z M, HUANG L J, WANG H M, ZHOU C Y. Analysis on differences in nutritional ingredients betweenand[J]. Quality and Safety of Agro-products, 2017(1): 73-78. (in Chinese)
[5] LI Y, SHENG Y, LU X, GUO X, DU P. Isolation and purification of acidic polysaccharides fromand evaluation of their lipid-lowering mechanism[J]. International Journal of Biological Macromolecules, 2020, 157: 276-287
[6] 黄在兴, 王义祥, 赵光辉, 翁伯琦, 杨丽萍, 刘朋虎. 姬松茸育种研究进展[J]. 亚热带农业研究, 2020, 16(4): 273-278.
HUANG Z X, WANG Y X, ZHAO G H, WENG B Q, YANG L P, LIU P H. Research progress onbreeding[J]. Subtropical Agriculture Research, 2020, 16 (4): 273-278. (in Chinese)
[7] LIU P H, YUAN J, JIANG Z H, WANG Y X, WENG B Q, LI G X. A lower cadmium accumulating strain ofproduced by60Co-γ-irradiation[J]. LWT-Food Science and Technology, 2019, 114: 108370.
[8] 鲍大鹏, 王 南, 谭 琦, 潘迎捷. 用荧光染色法对柱状田头菇()子实体担子和担孢子的观察[J]. 南京农业大学学报, 2000(3): 57-60.
BAO D P, WANG N, TAN Q, PAN Y J. Observation on basidia and basidiospores ofby fluorescence staining[J]. Journal of Nanjing Agricultural University, 2000(3): 57-60. (in Chinese)
[9] 许昭仪, 李 浩, 张 平. 金针菇生活史各阶段核相研究[J]. 菌物学报, 2015, 34(3): 386-393.
XU Z Y, LI H, ZHANG P. Behavior of nuclei in life cycle of[J]. Mycosystema, 2015, 34(3): 386-393. (in Chinese)
[10] 王瑞娟, 万佳宁, 龚 明, 茅文俊, 汪 莹, 唐利华, 尚俊军, 杨 慧, 鲍大鹏. 改良石蜡切片法观察金针菇子实体不同发育时期组织结构[J]. 食用菌学报, 2019, 26(2): 54-58.
WANG R J, WAN J N, GONG M, MAO W J, WANG Y, TANG L H, SHANG J J, YANG H, BAO D P. Tissue structure observation offruiting bodies at different developmental stages by improved paraffin method[J]. Acta Edulis Fungi, 2019, 26(2): 54-58. (in Chinese)
[11] 董金福. 竹林地仿生栽培姬松茸技术研究[D]. 杭州: 浙江农林大学, 2011.
DONG J F. The research of bionic cultivation technology ofin bamboo forest land[D]. Hangzhou: Zhejiang Agriculture and Forestry University, 2011. (in Chinese)
[12] 赵建霞, 冯伟林, 金群力, 沈颖越, 宋婷婷, 蔡为明. 双孢蘑菇担子与担孢子形态发育显微观测[J]. 园艺学报, 2019, 46(2): 356-364.
ZHAO J X, FENG W L, JIN Q L, SHEN Y Y, SONG T T, CAI W M. Microscopic observation of the morphological development of basidia and basidiospores in[J]. Acta Horticulturae Sinica, 2019, 46(2): 356-364. (in Chinese)
[13] 胡晓棣, 李 熠, 任蜀豫, 姚一建. 冬虫夏草、蛹虫草菌丝隔膜和细胞核荧光染色[J]. 菌物学报, 2016, 35(9): 1099-1105.
HU X D, LI Y, REN S Y, YAO Y J. Fluorescent staining of septa and nuclei inand[J]. Mycosystema, 2016, 35(9): 1099-1105. (in Chinese)
[14] HERRRIRA K M, ALVES E, COSTA M D, DIAS E S. Electron microscopy studies of basidios porogenesis in[J]. Mycologia, 2012, 104(6): 1272-1280
[15] 谭 伟, 周 洁, 曹雪莲, 苗人云, 贾定洪, 黄忠乾, 甘炳成, 彭卫红, 郑林用. 姬菇258和金凤2-1杂交构建姬菇新菌株[J]. 西南农业学报, 2015, 28(6): 2655-2660.
TAN W, ZHOU J, CAO X L, MIAO R Y, JIA D H, HUANG Z Q, GAN B C, PENG W H, ZHENG L Y. Hybrid strains breeding byNo.258 andsp Jinfeng2-1[J]. Southwest China Journal of Agricultural Sciences, 2015, 28(6): 2655-2660. (in Chinese)
[16] 于 浩, 葛志豪, 徐丽丽, 郭立忠. 双单杂交技术选育长根菇高品质抗病新菌株[J]. 南方农业学报, 2019, 50(12): 2621-2628.
YU H, GE Z H, XU L L, GUO L Z. Breeding of high oudenone production andresistantstrain by dikaryon monokaryon matin[J]. Journal of Southern Agriculture, 2019, 50(12): 2621-2628. (in Chinese)
[17] 任纪帆, 朱静娴, 王庆佶, 高馨梅, 姜淑霞. 大球盖菇“山农球盖3号”品种的选育[J]. 菌物学报, 2020, 39(6): 977-982.
REN J F, ZHU J X, WANG Q J, GAO X M, JIANG S X. Breeding of“Shannongqiugai No. 3”[J]. Mycosystema, 2020, 39(6): 977-982. (in Chinese)
[18] 陈美元, 廖剑华, 郭仲杰, 李洪荣, 卢政辉, 蔡丹凤, 王泽生. 双孢蘑菇耐热相关基因的表达载体构建及转化研究[J]. 菌物学报, 2009, 28(6): 797-801.
CHEN M Y, LIAO J H, GUO Z J, LI H R, LU Z H, CAI D F, WANG Z S. The expression vector construction and transformation of thermotolerance-related gene of[J]. Mycosystema, 2009, 28(6): 797-801. (in Chinese)
[19] 乔燕楠, 林星雨, 高 雅, 闫 静, 张岚岚, 李南羿. 根癌农杆菌介导的双孢蘑菇转基因体系的建立[J]. 食用菌学报, 2021, 28(1): 48-54.
QIAO Y N, LIN X Y, GAO Y, YAN J, ZHANG L L, LI N Y. Establishment of a high-efficiency agrobacterum mediated transformation system in[J]. Acta Edulis Fungi, 2021, 28(1): 48-54. (in Chinese)
[20] 喻义赣. 香菇农杆菌介导转化条件优化及BCA基因转化研究[D]. 福州: 福建农林大学, 2009.
YU Y G. Research of the agrobacterium-mediated transformation condition optimization and transformation of BCA gene in[D]. Fuzhou: Fujian Agriculture and Forestry University, 2009. (in Chinese)
[21] 杨利梅. 砖红绒盖牛肝菌子实体、菌丝和原基的显微及超微结构观察[D]. 昆明: 云南大学, 2017.
YANG L M. Microstructure and ultrastructure observation of fruit bodies, mycelia and primordia of[D]. Kunming: Yunnan University, 2017. (in Chinese)
[22] 郭 勇, 叶小金, 甘炳成, 彭卫红. 不同温度和光照培养条件下3种食用菌菌丝的菌落及菌丝形态的研究[J]. 西南农业学报, 2011, 24(6): 2301-2306.
GUO Y, YE X J, GAN B C, PENG W H. Study on three kinds of edible fungi hyphae colony and hyphae form in different temperature and light training conditions[J]. Southwest China Journal of Agricultural Sciences, 2011, 24(6): 2301-2306. (in Chinese)
[23] 徐思佳, 万佳宁, 李 焱, 李 燕, 尚俊军, 鲍大鹏. 刺芹侧耳交配型基因敲入单核体后锁状联合和核相的表征观察[J]. 菌物学报, 2020, 39(6): 1130-1138.
XU S J, WAN J N, LI Y, LI Y, SHANG J J, BAO D P. Characterization of clamps and nuclear phases in the monokaryons with transformed mating genes in[J]. Mycosystema, 2020, 39(6): 1130-1138. (in Chinese)
Microscopic Observations of Gills, Basidiospores and Mycelium in
ZHOU Yu1,2, HUANG Zaixing3, BAO Xinglu4, LI Jing2, LIU Penghu2*
1. College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China; 2. National Engineering Research Center of Juncao Technology, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China; 3. Institute of Applied Microbiology, College of Agriculture, Guangxi University / Institute of Edible Fungi, Nanning, Guangxi 530005, China; 4. Wutun Town Rural Revitalization Development Center, Wuyishan, Fujian 354309, China
is a precious edible and medicinal mushroom, which is not only rich in nutrition but also has high medicinal value, and its industrial development has a broad prospect. However, the lack of germplasm resources ofand insufficient knowledge of the genetic background ofhave hindered the development of its hybrid breeding technology, and the existing germplasm resources can hardly meet the practical needs of production.strains J1 was used as research materials,it is proposed to investigate the biological properties related toby micro-morphological observation of the reproduction-related structures of. The microscopic morphological characteristics of gills, basidiospores and mycelium in different growth and development stages were observed by scanning electron microscope and fluorescence microscope. The gills and trama were in the developmental stage and the hyphae lamellae were not obvious, and the gill sheets were not obvious during the morula state. The gill sheets were obviously observed,arranged parallel to each other,composed of trama and the fruiting layers on both sides, which were closely arranged, uniform in thickness, and a small amount of basidia were borne after the coral stage, but the trama was irregularly arranged. During the forming stage, the gills were clear and regular, and the inner veil was not ruptured, the gill space contains basidiospores and disorganized duct-like cells arranged in raised surface, and the duct-like cells in the gill were closely related. During the forming stage, the gill basidium was relatively full and wrinkled. There were 3 or 4 sporozoites on the gill basidia, and there were small connecting filaments between the basidiospores. The observation of basidiospore nuclei by DAPI staining showed that the most basidiospores were dinuclear spores, and a few were mononuclear and enucleated spores. The observation of mycelial nuclei by DAPI staining showed that the mycelium ofmycelium was multinucleated and had septa, but there was no lock-like association. This study would clarify the microscopic morphological characteristics of gills, basidia, basidiospores and mycelium in different growth and development stages of, enrich the knowledge of biological studies and life history processes of, and lay a foundation for the hybrid breeding, basidiospore mating types, genetics transformation system of
; gills; basidiospore; nucleus
S567.3
A
10.3969/j.issn.1000-2561.2022.12.010
2022-01-12;
2022-03-27
福建省科技厅农业引导性(重点)项目(No. 2020N0007);
福建农林大学学科交叉融合项目(No. XKJC-712021030);
中央引导地方科技发展专项(No. 2020L3030)。
周 宇(1997—),男,硕士,研究方向:食用菌栽培与育种。*通信作者(Corresponding author):刘朋虎(LIU Penghu),E-mail:phliu1982@163.com。
猜你喜欢 担子松茸菌丝体 自己的担子自己帮着担思维与智慧·上半月(2022年6期)2022-06-08响应面法优化鸡腿菇菌丝体多糖的提取工艺化学与生物工程(2021年1期)2021-01-22山东茌平:种植赤松茸让秸秆变废为宝今日农业(2020年23期)2020-12-15担子越重越要爱护挑担人当代陕西(2019年24期)2020-01-18一场直面问题、压实担子的大考人大建设(2019年11期)2019-05-21疯狂的虫草,疯狂的松茸和疯狂的岷江柏*——专访作家阿来阿来研究(2018年1期)2018-07-10如何保鲜一颗松茸?知识经济·中国直销(2018年6期)2018-06-29姬松茸当松茸卖是不是售假方圆(2016年24期)2017-01-16葡萄糖酸钠发酵废弃菌丝体提取壳聚糖的研究中国酿造(2016年12期)2016-03-01新型环保吸声材料——菌丝体胶合秸秆安全(2015年7期)2016-01-19猜你喜欢
- 2024-01-20 有关于第五次全国经济普查统计重点业务综合培训大会上讲话(完整文档)
- 2024-01-20 “严纪律、转作风、保安全、树形象”专题学习教育活动通知(完整文档)
- 2024-01-20 2024XX区住房城乡建设工作情况汇报
- 2024-01-20 2024高校思政教育交流材料:善用反腐败斗争这堂“大思政课”(精选文档)
- 2024-01-20 2024年主题教育专题党课辅导报告,(4)
- 2024-01-20 关于赴某地学习考察地方立法工作情况报告(范文推荐)
- 2024-01-20 2024年度关于增强党建带团建工作实效对策与建议(精选文档)
- 2024-01-20 教师演讲稿:春风化雨育桃李,,潜心耕耘满芬芳(全文)
- 2024-01-20 主题教育第二阶段来了
- 2024-01-20 2024年度关于到信访局实践锻炼个人总结【完整版】
- 搜索
-
- 打赌输了任人处理作文1000字7篇 05-12
- 当代大学生在全面建设社会主义现代化强 05-12
- 全面建成社会主义现代化强国的战略安排 03-10
- 个人廉洁自律方面存在的问题及整改措施 05-12
- 谈谈青年大学生在中国式现代化征程上的 05-12
- 2022年党支部第一议题会议记录(全文完 11-02
- 作为青年大学生如何肩负时代责任6篇 05-12
- 村党组织建设现状及工作亮点存在问题与 05-12
- 全面从严治党,自我革命重要论述研讨会 05-12
- 产业工人队伍建设改革(全文完整) 10-31
- 11-25国庆70周年庆典晚会 庆典晚会串词
- 11-25办公室礼仪的十大原则 浅谈办公室的电话礼仪
- 01-17用心灵轻轻地歌唱_心灵的歌唱
- 01-17也许你不是我一生的唯一|也许不是我
- 01-17爱了,请珍惜;不爱,趁早放手|爱就珍惜不爱就放手
- 01-17岁月带走的是记忆,但回忆会越来越清晰|有趣又有深意的句子
- 01-17曾经的美好只是曾经,我只想珍惜身边的人|我只想珍惜你
- 01-18从容不惊 [学会笑眼去看世界,不惊不乍,淡定从容]
- 02-03当代大学生学习态度调查报告
- 02-03常用护患英语会话
- 标签列表
