首页 > 心得体会 > 学习材料 / 正文
苹果愈伤组织基因编辑技术的建立
2023-02-05 15:10:08 ℃董超华,于家惠,刘瑛双,王南,曲衍杰,孙欣,柏素花,张玉刚
(1.青岛农业大学生命科学学院,山东青岛 266109;
2.青岛农业大学园艺学院/青岛市园艺植物遗传改良与育种重点实验室;
3.山东省高校植物生物技术重点实验室,山东青岛 266109)
基因编辑技术是一种能对生物基因组特定序列进行精确修改的基因工程技术,是21世纪最重要的生物技术之一。基因编辑技术主要依赖4种核酸内切酶,包括归巢核酸内切酶(meganuclease)、锌指核酸酶(zinc finger nucleases,ZFNs)、类转录激活因子效应物核酸酶(transcription activator-like effector nucleases,TALENs)和成簇规律间隔短回文重复序列(clustered regulatory interspaced short palindromic repeat,CRISPR)系统。
归巢核酸内切酶能够特异识别12~40 bp的DNA序列,造成DNA双链断裂,细胞在修复断裂的DNA时,会在断裂位点随机插入或删除核苷酸[1]。ZFNs是将限制性核酸内切酶FokⅠ[2]切割域与锌指模块融合而成,通过改造锌指模块可靶向切割DNA序列,造成双链断裂。但该技术效率低、成本高、易脱靶。TALENs与ZFNs的形式类似,是由FokⅠ内切酶切割域与TALEN蛋白DNA结合域融合组成。与前两种技术相比,TALENs特异性较高[3],但模块组装过程繁琐,具有一定的细胞毒性。CRISPR/Cas系统由CRISPR序列元件和CRISPR相关蛋白(CRISPR-associated proteins,Cas)基因家族组成,CRISPR包含一个crRNA(CRISPR RNA)、一个反式激活crRNA(tracrRNA)和Cas9蛋白。其中crRNA用于识别靶序列,tracrRNA用于促进crRNA成熟,Cas9蛋白用于切割靶序列。为方便应用,将tracrRNA和crRNA融合在一起形成单一引导RNA(sgRNA)。前三种内切酶的共同特点是:直接与靶序列DNA结合并切割,不依赖同源识别,而CRISPR/Cas9系统依赖sgRNA识别靶序列,然后由Cas9进行切割。这几种核酸内切酶都能造成双链断裂,断裂后由细胞通过两种修复途径进行修复:同源重组(homologous recombination,HR)和非同源端连接(nonhomologous end joining,NHEJ)[4]。在连接过程中造成随机少量核苷酸的插入或缺失,使基因的编码区发生移框,从而影响基因的表达导致功能丧失。
CRISPR/Cas9系统因其简单、精准、高效的特点被广泛应用于动植物基因功能的挖掘及定向改良。例如基因编辑技术推动了转基因动物的发展,基因编辑小鼠[5-6]、基因编辑猴[7]和基因编辑猪[8]成为人类疾病建模的主要工具和器官移植的潜在器官供体,对人类疾病研究有重要贡献。植物上,基因编辑在调控株型[9]、增强抗病性[10-11]、提高产量[12]等方面有广泛的研究和应用。苹果作为我国重要的经济作物,基因编辑技术对其遗传改良具有重要意义,可用于改善抗病性[13],获得提早开花的突变植株[14],调控果实营养物质的合成[15]等。但受限于其极低的转化效率,苹果的基因编辑技术进展很慢,还远不成熟,本文以苹果愈伤组织为试验材料,通过改造载体、靶序列验证,获得编辑成功的苹果愈伤组织,初步建立基于CRISPR/Cas9系统的基因编辑技术。
1.1 材料
1.1.1 试验材料
试验材料包括苹果(Malusdomestica)‘Orin’愈伤组织和本生烟(Nicotinanabenthamiana)。‘Orin’愈伤组织在Y1培养基(4.4 g/L MS,0.225 mg/L 6-BA,30 g/L蔗糖,8 g/L琼脂,1 mg/L 2,4-D,pH 5.8)上,25 ℃暗培养。本生烟培养在人工气候箱内,25 ℃、8 h/16 h的光暗循环。
1.1.2 试验药品
MS(Murashige &Skoog Medium)培养基,广泛应用于植物器官、花药、细胞和原生质体的培养;
6-苄氨基腺嘌呤(6-Benzylaminopurine,6-BA),细胞分裂素;
2,4-二氯苯氧乙酸(2,4-Dichlorophenoxyacetic acid,2,4-D),具有代表性的合成植物生长素(auxin);
萘乙酸(1-Naphthaleneacetic acid,NAA),促进植物根系生长的植物生长调节剂;
2-(N-吗啉代)乙烷磺酸一水(MES monohydrate,MES),用于缓冲液;
乙酰丁香酮(Acetosyringone,AS),可诱导农杆菌Vir基因的活化,从而促进外源基因的整合。
1.2 试验方法
1.2.1 载体的构建
根据CCTop-CRISPR/Cas9 target online predictor 在线(http://crispr.cos.uni-heidelberg.de/)预测目标基因的编辑靶位点。选定的靶序列与基因组序列进行比对,以确保靶位点的连续性,排除跨越内含子的靶点序列。本试验对MdVPE1进行编辑,用CCTop选定的靶点序列为5′-TCTTTTTCCCTCATCGGCAGTGG-3′和5′-CAAGTAATCCTACAGAACCATGG-3′。选用的载体是用于拟南芥的基因编辑载体pHDE-35S-Cas9-mCherry-UBQ[16]。为方便苹果愈伤组织选择,首先将载体原有的筛选潮霉素抗性基因HYG换成卡那霉素抗性基因Kan,并利用同源重组将gRNA和AtU6-26终止子连接到载体上。随后,利用PCR将MdU6启动子和基因靶位点序列进行整合,将PCR产物回收后,通过同源重组插入到gRNA序列前。将构建完成并测序正确的质粒转化农杆菌EHA105菌株,用于后续愈伤组织的遗传转化。
1.2.2 本生烟叶片的瞬时转化
将农杆菌菌液培养至OD600=0.8,倒入50 mL离心管中,5 500 r/min 离心10 min,弃上清液,将沉淀用同体积的侵染液重悬(10 mmol/L MgCl2·6H2O、10 mmol/L MES、0.1 mmol/L AS,pH 5.8)。选取生长状态良好的4~6周本生烟草叶片,用1 mL 注射器吸取重悬后的菌液,在烟草叶片背面用压力渗入法将农杆菌注入叶片中。
1.2.3 愈伤组织的遗传转化
取转化后的农杆菌菌液100 μL加入到含有相应抗生素的50 mL LB液体培养基中,于28 ℃,200 r/min摇床中培养农杆菌至OD600=0.8。将新鲜的菌液转入50 mL离心管,5 500 r/min离心10 min,收集菌体;
用等体积愈伤组织侵染液RM(4.4 g/L MS、30 g/L蔗糖、100 μmol/L AS)重悬,28 ℃振荡培养30 min;
在超净工作台中用高温灭菌后的镊子挑取20 d的愈伤组织移至重悬后的菌液中,并将较大块的愈伤组织捣碎,侵染15 min;
将侵染后的愈伤组织用无菌纱布过滤,并用滤纸吸干菌液;
将愈伤组织均匀铺到共培养基Y1上,黑暗中25 ℃共培养2~3 d。
共培养后,将愈伤组织转移至含有100 mg/L的Kan抗生素的筛选培养基Y2(4.4 g/L MS、0.225 mg/L 6-BA、30 g/L蔗糖、8 g/L琼脂、1 mg/L萘乙酸,pH 5.8,灭菌后加入100 mg/L Kan)上,25 ℃暗培养,直至长出阳性愈伤组织。转移阳性愈伤组织至新鲜Y2培养基上继代培养,并提取DNA,进行鉴定。
1.2.4 基因编辑结果的鉴定
根据靶位点前后各300 bp左右的基因组DNA序列设计特异性引物,提取阳性愈伤组织DNA,并以其为模板用高保真酶进行PCR扩增,将扩增产物连接到pMD18-T载体上。挑取单菌落进行菌液PCR筛选后,测序,根据测序结果确定靶位点是否编辑成功,如发现靶序列发生改变,则用二代测序技术进行扩增子测序。
2.1 MdU6启动子的克隆
以拟南芥U6-26基因为电子探针,用一套在蛋白质数据库或DNA数据库中进行相似性比较的分析工具 (basic local alignment search tool,BLAST)在苹果基因组数据库中分析其同源序列,获得其同源基因序列(命名为MdU6)。根据基因编码区上游1 kb 基因组 DNA序列设计引物,以苹果基因组DNA为模板扩增获得其编码区上游序列。将获得的序列与拟南芥U6-26启动子序列进行比对,确定苹果MdU6基因启动子(图1),注册序列号为MT584802。

图1 苹果MdU6启动子和拟南芥U6-26启动子序列比对Fig.1 Sequence alignment between apple U6 promoter and Arabidopsis U6-26 promoter注:黑色背景表示一致的核苷酸,灰色背景表示相似的核苷酸。
2.2 靶序列的验证
为验证目标基因靶序列的有效性,将目标基因的靶点序列及间隔序列毗邻基序(protospacer adjacent motif,PAM)整合到绿色萤光蛋白(green fluorescent protein,GFP)基因的5′端构建植物表达载体,与基因编辑载体分别转入农杆菌,然后将含有两种载体的农杆菌注射到本生烟叶片中,使它们在烟草叶片中共表达,并进行基因编辑。同时,对上述整合到GFP基因上的PAM位点进行点突变以破坏PAM位点,并以新的载体与基因编辑载体共表达作为对照,比较对照和编辑部位荧光强度。结果显示,MdVPE1编辑载体与PAM位点突变的GFP融合表达载体的农杆菌共转烟草后的荧光强度明显高于MdVPE1编辑载体与GFP融合表达载体的农杆菌共转烟草后的荧光强度,而单一MdVPE1编辑载体的农杆菌转烟草后,无荧光产生(图2)。

图2 基因编辑靶点序列有效性的快速验证Fig.2 Rapid verification of the effectiveness of gene editing target sequences注:图中所示为本生烟叶片注射农杆菌后2 d观察的GFP荧光;
红色虚线所示为注射区域,图中文字说明响应区域所注射农杆菌中包含的载体。
2.3 CRISPR载体的构建及愈伤组织的遗传转化
为探索基因编辑在苹果愈伤组织中的有效性,将苹果液泡加工酶基因(MdVPE1)作为目标基因构建基因编辑载体。液泡加工酶是植物体内参与植物抗病性的一种蛋白酶,该酶突变会使植物抗性发生改变。利用在线软件CCTop选择靠近基因编码区5′端的靶位点,并与苹果基因组数据库进行比对,保证其特异性。基因编辑载体框架采用pHDE-35S-Cas9-mCherry-UBQ质粒。采用卡那霉素抗性基因作为报告基因。为提高转录效率,以苹果自身的MdU6启动子启动gRNA的转录,采用拟南芥U6-26终止子。按苹果的密码子偏好优化了Cas9蛋白的编码序列(图3)。

图3 苹果MdVPE1基因编辑载体的T-DNA区域组成Fig.3 Composition of T-DNA region of apple MdVPE1 gene editing vector
利用农杆菌介导法,将上述构建完成的基因编辑载体转化‘Orin’愈伤组织,以验证基因功能。侵染后,在Y1培养基上共培养3 d,移至含有Kan抗生素的培养基进行抗性筛选,待长出新的愈伤凸起,移至继代培养基中继续继代培养(图4)。

a.愈伤组织在筛选培养基上共培养;
b.愈伤组织在筛选培养基上长出凸起的抗性愈伤组织。图4 愈伤组织的遗传转化Fig.4 Genetic transformation of apple callus
2.4 苹果MdVPE1基因编辑结果鉴定
将测序结果与原基因序列比对后发现,靶位点1和靶位点2均存在多种突变类型的混合突变体。其中,靶位点1分别出现了8个碱基的缺失、13个碱基的缺失。靶位点2分别出现了8个碱基的缺失、4个碱基的替换和5个碱基的插入。这些突变使蛋白质编码区(sequence coding for amino acids in protein,CDS)序列编码紊乱,导致MdVPE1基因的敲除及功能丧失(图5)。

a.MdVPE1靶位点位置示意图;
b.MdVPE1基因突变测序结果示意图(红色代表PAM位点,蓝色代表突变位点,绿色代表插入位点);
c.MdVPE1基因突变测序结果峰图。图5 苹果愈伤组织MdVPE1基因编辑后的2个编辑位点基因组测序结果出现6种突变Fig.5 Six mutations in genome sequencing results of 2 editing sites after MdVPE1 gene editing in apple callus
2.5 MdVPE1基因编辑愈伤组织对轮纹病菌的抗性分析
为了进一步确定MdVPE1基因编辑后是否影响苹果愈伤组织的抗病性,对基因编辑发生突变的愈伤组织(knock out,KO)、转化空的CRISPR载体作为对照愈伤组织(empty vector,EV)和野生型愈伤组织(wild type,WT)分别接种病原菌进行鉴定。用继代3周的KO、EV和WT愈伤组织进行接种试验,这3种愈伤组织没有明显的表观差异。在KO、EV和WT愈伤组织上分别接种轮纹病病原菌(B.dothidea)。以菌丝延伸区域直径作为评估愈伤组织抗性的指标,接种5 d后,真菌侵染区域在KO和EV之间或KO和WT之间表现出极显著差异(p<0.01,图6)。与EV和WT愈伤组织相比,KO愈伤组织的菌丝显著减少,延伸速度较慢,表明基因编辑导致愈伤组织对B.dothidea的抗性发生改变。
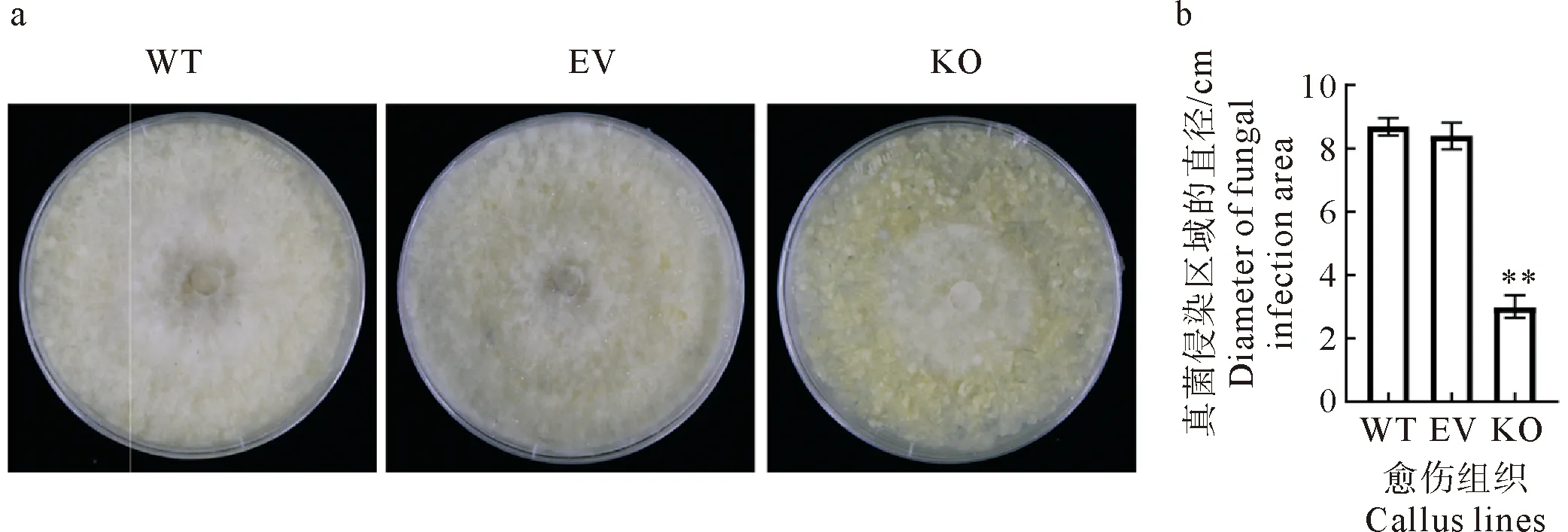
a.病原菌在不同的愈伤组织中表现不同的生长速度;
b.病原菌的延伸区域直径在基因编辑愈伤组织中显著小于野生型和空载体对照,** 代表在p<0.01的水平上差异极显著。图6 MdVPE1 基因编辑导致愈伤组织抗病性发生改变Fig.6 Changes in callus disease resistance caused by MdVPE1 gene editing
目前,CRISPR/Cas9基因编辑技术已在拟南芥、番茄、本生烟等模式植物中应用广泛。与传统的物理突变不同,利用该技术可以对靶基因的特定位点进行编辑,实现精准的基因突变。通过本生烟草瞬时表达进行靶序列验证,结果发现,MdVPE1编辑载体与PAM位点突变的GFP融合表达载体的农杆菌共转烟草后的荧光强度明显高于MdVPE1编辑载体与GFP融合表达载体的农杆菌共转烟草后的荧光强度,而单独的MdVPE1编辑载体的农杆菌瞬转烟草后无荧光。这表明PAM位点突变后,编辑载体无法识别NGG结构,未能对靶位点进行基因编辑,从而无法影响靶位点下游的GFP的表达;
而当PAM位点未发生突变时,编辑成功,使CDS 序列编码紊乱,导致基因发生突变,影响GFP的正常表达,表现出荧光强度的大幅度减弱。利用这一方法可以快速筛选高效的靶序列。
本文利用CRISPR/Cas9技术成功编辑苹果愈伤组织中MdVPE1基因。对获得的阳性愈伤组织进行DNA测序,结果发现愈伤组织中MdVPE1基因的CDS序列在靶位点均出现碱基突变。且同一愈伤组织中出现不同类型的碱基突变,这表明我们所获得的阳性愈伤组织不是单细胞的克隆,或者这些细胞中的编辑行为不是一次性完成的。统计分析突变类型发现,主要包括非3整数倍数的多个碱基的缺失、不同数目碱基的插入及替换。这些突变使基因CDS序列中断,最终导致愈伤组织中MdVPE1基因的敲除及功能丧失,表明成功实现基因编辑。分析MdVPE1基因编辑愈伤组织对轮纹病菌的抗性发现,接种5 d后,与EV和WT愈伤组织相比,KO愈伤组织的菌丝显著减少,表明基因编辑导致愈伤组织对B.dothidea的抗性发生改变。
本文利用 CRISPR/Cas9 技术成功敲除苹果愈伤组织中MdVPE1基因,建立了基于 CRISPR/Cas9 系统的基因编辑技术,为研究苹果基因功能奠定了重要基础。
猜你喜欢 抗性位点测序 Pd改性多活性位点催化剂NH3-SCR脱硝反应机理研究分子催化(2022年1期)2022-11-02两种高通量测序平台应用于不同SARS-CoV-2变异株的对比研究中国人兽共患病学报(2022年9期)2022-10-19不忘初心,回归经典!养虾至暗时刻,海茂坚定高抗苗,携抗性更强“普利茂”回来了当代水产(2022年7期)2022-09-20冻干益生菌微胶囊保护剂及抗性研究中国饲料(2021年17期)2021-11-02维生素D受体基因Bg1I、Cdx-2位点多态性与桥本氏甲状腺炎的相关性昆明医科大学学报(2021年8期)2021-08-13基于网络公开测序数据的K326烟草线粒体基因组RNA编辑位点的鉴定与分析烟草科技(2021年6期)2021-06-24生物测序走在前科学导报(2021年29期)2021-06-03外显子组测序助力产前诊断胎儿骨骼发育不良中国生殖健康(2020年4期)2021-01-18MEAM1烟粉虱隐种对3种新烟碱类杀虫剂交互抗性及交互现实遗传力的分析新疆农业科学(2020年9期)2020-10-14基因测序技术研究进展科海故事博览·下旬刊(2019年6期)2019-04-16猜你喜欢
- 2024-01-20 有关于第五次全国经济普查统计重点业务综合培训大会上讲话(完整文档)
- 2024-01-20 “严纪律、转作风、保安全、树形象”专题学习教育活动通知(完整文档)
- 2024-01-20 2024XX区住房城乡建设工作情况汇报
- 2024-01-20 2024高校思政教育交流材料:善用反腐败斗争这堂“大思政课”(精选文档)
- 2024-01-20 2024年主题教育专题党课辅导报告,(4)
- 2024-01-20 关于赴某地学习考察地方立法工作情况报告(范文推荐)
- 2024-01-20 2024年度关于增强党建带团建工作实效对策与建议(精选文档)
- 2024-01-20 教师演讲稿:春风化雨育桃李,,潜心耕耘满芬芳(全文)
- 2024-01-20 主题教育第二阶段来了
- 2024-01-20 2024年度关于到信访局实践锻炼个人总结【完整版】
- 搜索
-
- 打赌输了任人处理作文1000字7篇 05-12
- 当代大学生在全面建设社会主义现代化强 05-12
- 全面建成社会主义现代化强国的战略安排 03-10
- 个人廉洁自律方面存在的问题及整改措施 05-12
- 谈谈青年大学生在中国式现代化征程上的 05-12
- 2022年党支部第一议题会议记录(全文完 11-02
- 作为青年大学生如何肩负时代责任6篇 05-12
- 村党组织建设现状及工作亮点存在问题与 05-12
- 全面从严治党,自我革命重要论述研讨会 05-12
- 产业工人队伍建设改革(全文完整) 10-31
- 11-25国庆70周年庆典晚会 庆典晚会串词
- 11-25办公室礼仪的十大原则 浅谈办公室的电话礼仪
- 01-17用心灵轻轻地歌唱_心灵的歌唱
- 01-17也许你不是我一生的唯一|也许不是我
- 01-17爱了,请珍惜;不爱,趁早放手|爱就珍惜不爱就放手
- 01-17岁月带走的是记忆,但回忆会越来越清晰|有趣又有深意的句子
- 01-17曾经的美好只是曾经,我只想珍惜身边的人|我只想珍惜你
- 01-18从容不惊 [学会笑眼去看世界,不惊不乍,淡定从容]
- 02-03当代大学生学习态度调查报告
- 02-03常用护患英语会话
- 标签列表
